Не секрет, что самым большим органом человеческого тела является его кожа. Помимо защиты тела от внешних раздражителей, кожа выполняет еще и функцию датчика, собирающего информацию, наряду с глазами, ушами, языком и носом. Информация, получаемая кожей, позволяет человеку оценивать окружающую среду, лучше понимать ситуацию, в которой он находится и действовать в соответствии с ней. Несмотря на огромную важность тактильной информации, о том как именно все работает мы пока знаем не особо много. Посему ученые из Калифорнийского университета (США) решили рассмотреть кожу человека под математическим углом, дабы понять механизм возникновения и передачи тактильных ощущений. Что происходит, когда мы берем что-то в руки, как наша кожа обрабатывает получаемую информацию, и как данное исследование применить на практике? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
У взрослого человека площадь его кожи может достигать 2.3 м2, что делает ее самым большим органом. Однако габариты ничто, если за ними нет никакого функционала. Кожа выполняет достаточно много функций: защитная, дыхательная, экскреторная, терморегуляторная, иммунная, метаболическая и т.д.
Другими словами, пытаясь оценивать разные органы по их важности, ставить кожу на последнее место было бы ошибочно. Самой же загадочной функцией кожи является сбор информации, т.е. формирование осязания — одного из видов чувств человека. Такова температура в комнате, шершавые или гладкие обои, насколько мягкое кресло — все эти и многие другие данные собираются именно кожей.
Невероятная чувствительность кожи заключается в наличии колоссального числа нервных окончаний, т.е. рецепторов. Все они отличаются друг от друга по форме и строению, поскольку выполняют разные задачи (одни собирают информацию про фактуру объекта, другие — про температуру, например).
Рецепторы кожи можно разделить на два основных типа: свободные нервные окончания и несвободные нервные окончания. Первые состоят исключительно из конечных ветвлений осевого цилиндра и располагаются в эпителии. Эти рецепторы собирают данные о температуре (терморецепторы), давлении (механорецепторы) и болевых ощущениях (ноцицепторы). Категоризация несвободных нервных окончаний куда более обширная:
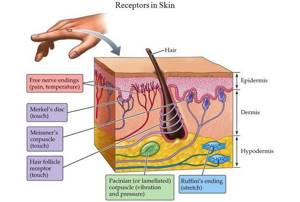 Категоризация несвободных нервных окончаний куда более обширная:
Категоризация несвободных нервных окончаний куда более обширная: - тельца Пачини — рецепторы давления в подкожно-жировой клетчатке;
- тельца Мейснера — рецепторы давления в дерме;
- тельца Меркеля — рецепторы давления в глубоких слоях эпидермиса;
- тельца Руффини — рецепторы растяжения, реагирующие на тепло;
- колбы Краузе — рецепторы в надсосочковом слое дермы (якобы реагируют на холод, но это под вопросом);
- рецепторы волосяных фолликулов — механорецепторы, которые реагируют на изменение положения волоса.
Это лишь краткий перечень, без глубокого рассмотрения рецепторов, их функций и строения, но и этого достаточно, чтобы понять всю сложность кожи как органа чувств.
Сами исследователи трактуют осязание как кодирование механических сигналов, собранных кожей и подкожными тканями, в нейронные сигналы. Нейронные ответы на тактильные раздражители часто связаны с механическими воздействиями, возникающими из небольших участков кожи, однако есть свидетельства о том, что динамическое прикосновение вызывает механические волны в тактильном диапазоне частот, которые распространяются по всей руке, с переходными возбуждениями, затухающими в течение 30 мс. Таким образом, динамические тактильные воздействия могут стимулировать широкое распространение афферентации*.
Афферентация* — непрерывный поток нервных импульсов от органов чувств к нервной системе.Было обнаружено, что эти волны, вызванные прикосновением, способствуют тонкому восприятию и могут использоваться для определения характеристик объекта, к которому дотронулись, области контакта объекта с рукой и дальнейших действий. Также есть данные, что рецептивные поля нейронов в соматосенсорных областях коры мозга охватывают большие участки рук и нескольких пальцев. Большая площадь контакта на ранних этапах обработки сигналов побуждает корковые нейроны отвечать на входные сигналы, которые доставляются обратно в область контакта. Таким образом, соматосенсорная обработка может зависеть от информации, переносимой механическими волнами, которые распространяются в тканях в отдаленные участки, удаленные от мест непосредственного механического контакта. Ученые считают, если перенос механических волн в руке способствует эффективному кодированию соматосенсорной информации, то должна быть возможность описать тактильные стимулы в малых участках посредством информативных параметров. Другими словами преобразовать ощущение прикосновения в цифры. В своем труде ученые показывают, как механические волны в руке производят эффективное кодирование тактильных входных данных. Проведя опыты с использованием высокоточных датчиков, ученые смогли создать своего рода словарик пространственно-временных сигналов, которые в совокупности позволяют классифицировать входящую информацию с точностью более 95%. То есть им удалось создать карту, показывающую где и какие области кожи руки активируются при контакте с тем или иным объектом.
Результаты исследования
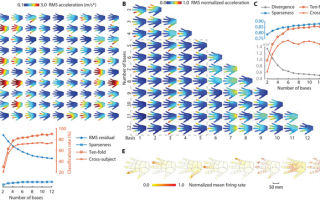
Моделирование тактильной информации ученые изобразили в виде матричного разложения. Оценка кодирования была выполнена посредством собранной в ходе опытов базы данных тактильных стимулов для всей кисти, включающую пространственно-временные изменения кожи a(x, t).
На руку добровольца были прикреплены специальные датчики в 30 участках (х). В ходе эксперимента было выполнено 13 жестов и 4600 взаимодействий с различными объектами. Изображение №1
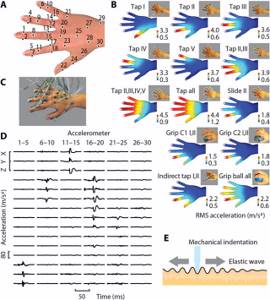 Изображение №1
Изображение №1 Каждый из стимулов wi(x, t), внесенный в набор данных, имел собственное время активации hi(t), которое также было учтено в модели для получения более точных «тактильных базовых паттернов» (2А), которые в совокупности кодируют все возникающие стимулы и передающиеся сигналы.
Изображение №2 Эти базисные паттерны (далее базисы) также могут быть интерпретированы как набор фильтров анализа, которые извлекают информацию из внешних стимулов с помощью различных дополнительных паттернов пространственно-временной интеграции механических сигналов в руке. По словам ученых, эти фильтры можно сравнить с функциями спектрально-временной настройки в слуховой обработке или с фильтрами пространственно-временного рецептивного поля при работе сетчатки. Суммируя, учеными была создана математическая модель, в которой сигналы, ощущаемые по всей руке, были представлены в виде небольшого числа упрощенных паттернов. Данная методика позволила получить основные волновые паттерны — вибрации кожи по всей кисти, которые участвуют в сборе и передаче тактильной информации.
Несмотря на то, что в анализе не учитывались условия возникновения сигналов, тактильные базисы напоминали сенсорную функцию кисти (2А и 2В). Большинство из них первоначально были локализованы на дистальных концах одного из пальцев (наиболее плотно иннервируемые области кисти).
Скорость движения сигналов составляла порядка 1-10 м/с, а затухание сигнала наблюдалось спустя 10-30 мс после его возникновения. Другие тактильные базисы эволюционировали от дистальной области отдельных пальцев до диффузных областей поверхности кисти (2А).
В аспекте частоты, пара базисов демонстрировала схожее пространственное расположение, но разные частотные характеристики.
К примеру, есть пара базисов, локализованных в пределах одного пальца, но имеющих разные фильтрационные свойства (относительно передаваемых сигналов): нижний диапазон от 20 до 80 Гц (2В, базис 2) или верхний диапазон от 80 до 160 Гц (2B, базис 6).
Изображение №3 Ученые считают, что пространственно-временные тактильные базисы связаны с определенным пальцем, т.е. имеют свою рабочую зону, так сказать. Например, 45% из 4600 проанализированных тактильных раздражителей были вызваны жестами, когда с объектом контактировал только один палец. Проведя повторный анализ, исключающий тактильные сигналы, создаваемые одним лишь пальцем, была обнаружена такая же тенденция.
Пространство возможных тактильных раздражителей ограничено механикой и продолжительностью контакта (3А).
Далее ученые решили проверить, сколько базисов должно быть задействовано для определения источника сигнала. Как оказалось, если использовать не менее 7, то точность определения составит 90%, а если 12, то 95%. Тем не менее, не все стимулы требуют активации столь большого числа базисов для повышения точности. Логика достаточно прямолинейна: когда в жесте задействовано несколько пальцев, то активируются несколько базисов; если же в жесте задействован лишь один палец, то и базисов будет один, максимум два. При этом сами базисы также варьировались в зависимости от жестов. То есть, разные жесты, хоть в них и задействованы одинаковые пальцы, будут активировать разные базисы.
Модель также показала, что достаточно пяти базисов для максимизации точности (80%), с которой стимулы от одного участника опытов могли быть классифицированы с использованием данных от других участников (3C). Эти пять базисов были практически универсальны среди всех участников и соответствовали пяти пальцам кисти (3B).
Совокупность вышеописанных наблюдений говорит о том, что сама эластичность кожи играет важную роль в сборе и передаче информации, поскольку за счет нее увеличивается площадь контакта с объектом. Кроме того, волны сигналов, распространяющиеся по определенному паттерну, позволяют классифицировать полученную информацию, что также способствует ускорению ее обработки непосредственно мозгом. Подобные механизмы обработки сигналов можно сравнить с работой среднего уха, которое распространяя звуки с различным частотным содержанием на разные сенсорные рецепторы в ухе, помогает кодированию звуков слуховой системой.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Данное исследование показало нам, что кожа является намного более сложной системой, чем считалось ранее. Если раньше процесс передачи сигналов можно было описать линейно (прикосновение — возникновение сигнала — передача сигнала в мозг), то сейчас этот процесс скорее похож на волновую активность.
Сигналы, получаемые от объектов взаимодействия с кожей, распространяются волнами по нервным окончаниям кожи в зависимости от зоны контакта, его продолжительности и характера поверхности. Другими словами, в сборе информации про объект контакта участвуют не только рецепторы в непосредственно месте контакта, но и рецепторы вокруг этой зоны.
Исследователи считают, что в этом сложном процессе не последнюю роль играет эластичность кожи, позволяющая увеличить площадь контакта с точки зрения распространения сигналов, а не с точки зрения непосредственно самого контакта.
По мнению ученых, их труд позволит не только лучше понять работу мозга и нервной системы человека, но и пригодится в разработке новых протезов и даже роботов, способных тактильно более точно собирать информацию об окружающей среде.
Пятничный офф-топ: Для создания протеза может потребоваться множество сложных запчастей и лабораторный комплекс или конструктор LEGO и креативный подход. Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! 🙂
Немного рекламы 🙂
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.
99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.
2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
Материалы для студентов МИРЭА — Механорецепция
- Доклад по биофизике:
- “Механорецепция”
- 1. Назначение
Для активного существования в окружающей среде, высшие организмы обладают большим количеством анализаторов, специализирующихся на разных типах воздействия: на свете (зрение), на звуке (слух), на вкус и запах (обаняние), на прикосновения и температуру (осязание), на гравитацию. Механорецепция входит в чувство осязания и вместе с вестибулярным аппаратам позволяет наиболее полно представить организму и сознанию положение конечностей и тела в пространстве.
- 2. Функции
- Анализатор механорецепции, как и любой другой анализатор, включает три пространственно-функциональных части: рецептор (получение и трансформация механического воздействия в электрический импульс), проводниковые пути (передача импульса) и нервный центр (анализ полученной информации и формирование эфферентного ответа). Все эти части обеспечивают:
— Восприятие механических воздействий на кожу: локализация, направление движения, скорость деформирующего источника, его вибрации (тактильная рецепция); — Восприятие механических сдвигов внутри органов и мышц с целью определения положения конечностей и тела в пространстве (проприоцепция); — Восприятие воздействий на волосяной покров. — Трансформацию сигнала из механического в электрический, который может быть передать по нейронам. — Передачу импульсов с высокой или низкой скоростью к нервному центру. — Формирование в сознании общей картины положения тела и конечностей в пространстве.
— Обеспечение вегетативной нервной системы информации о положении тела и его контроль (поддержание вертикального положения тела при возможном засыпании в таком положении, информация о глазодвигательных движениях во сне).
3. Устройство и принцип работы.
Типичные механорецепторы, как правило, представляют собой инкапсулированные образования. Некоторые из них называют поверхностными концевыми органами, т. к. они в коже расположены поверхностно. Это диски Меркеля, тельца Мейснера, тельца Фатера-Пачини, тельца Догеля, колбы Краузе, тельца Руффини, нервно-сухожильные веретена, нервно-мышечные веретена и прочие (Рис.1).
Рис.1. Различные виды механорецепторов[4]
Пластинчатые тельца (по старой терминологии тельца Фатера-Пачини) располагаются в соединительной ткани внутренних органов и глубоких слоях кожи, особенно на подушечках пальцев, на брыжейке, в молочной железе, кишечнике и других внутренних органах. Имеют вид округлых образований. Структурными компонентами являются:
— внутренняя колба (луковица), образованная видоизмененными леммоцитами, в которую проникают нервные волокна; — наружная колба – слоистая соединительнотканная капсула из фибропластов и коллагеновых волокон, образующих концентрические пластины, между которыми имеется жидкость.[2]
Внутренняя часть капсулы содержит плоские, концентрически расположенные нейроглиальные клетки, ограничивающие внутреннюю колбу. Рецепторное волокно входит во внутреннюю колбу с одного его полюса и образует контакты с глиальными клетками. Терминальная часть рецепторного волокна содержит мелкие сферические митохондрии и светлые синаптические пузырьки.
Внешний слой капсулы состоит из мощной соединительнотканной оболочки, образованной из плоских серповидных клеток и соединительнотканных волокон, между которыми находится интерстициальная жидкость. Пластинчатые тельца воспринимают ощущение вибрации, натяжения, давления на органы и внутриорганное давление.
Принцип работы телец Фатера-Пачини на сегодня плохо изучен. Соединительнотканные пластинки и интерстициальная жидкость капсулы вероятно способствуют усилению давления на нервное окончание, в результате чего аксолемма деформируется, изменяется её проницаемость и происходит генерация потенциала[1].
Полагают, что местом возникновения потенциала действия в тельцах Пачини является область первого перехвата Ранвье[2].
Осязательные тельца (Мейснера) размещаются в сосочковом слое дермы, имеют эллипсоидную форму и небольшие размеры. Это тактильные рецепторы, реагирующие на прикосновение. Присутствует в дерме кожи, особенно часто в кончиках пальцев, подошвах, сосках, веках, губах и половых органах.
В центре тельца Мейснера располагается спирально свёрнутое безмиелиновое разветвление миелинового волокна, которое проходит через поперечно расположенные овальные клетки, напоминающие шванновские клетки. Снаружи тельце покрыто соединительнотканной капсулой.
Внутренняя колба состоит из глиальных клеток, лежащих перпендикулярно длинной оси тельца, между которыми располагаются веточки дендритов. Снаружи находится очень тонкая, переходящая в периневрит слоистая капсула – наружная колба.
Незначительная деформация капсулы передается глиоцитам и далее на дендриты. [2]
Клетки Меркеля расположены под эпидермисом, обладают крупными неправильной формы ядрами и микроворсинками, простирающимися к эпидермальным клеткам.
У их оснований располагаются дисковидные окончания сенсорных аксонов (диски Меркеля). Группа из 10 — 20 клеток Меркеля образует синаптические контакты с окончаниями одного сенсорного аксона (тельца Пинкуса-Игго).
Клетки Меркеля реагируют на внезапные смещения кожи, например при поглаживании. [4]
Тельца Руффини лежат в соединительнотканной части кожи и капсулах суставов: они воспринимают давление и имеют вид веретеновидных структур. Внутреннюю колбу образуют глиальные клетки между которыми располагаются терминали дендритов с расширениями на концах. Капсула хорошо выражена.
Колбы Краузе – мелкие округлые тельца, являющиеся механорецепторами и холодовыми рецепторами. Они лежат в дерме кожи, слизистой оболочке полости рта, подгортанника, в конъюнктиве глаза. Внутренняя колба образована плоскими глиоцитами между которыми тонкие ветви дендрита образуют сплетения в виде клубочка. Наружная капсула очень тонкая.
Генитальные тельца Догеля – находятся в особо чувствительных областях кожи (наружные половые органы, молочные железы). Они по строению похожи на колбы Краузе, но в отличие от них в тельце входит несколько отростков от нейроцитов, что вызывает сильную иррадиацию возбуждения. Реагируют на давление импульсами, вызывающими половое возбуждение.[2]
Механочувствительные свободные окончания в коже. Являются немиелинизированными аксонами, у них нет корпускулярных структур. К ним относятся рецепторы волосяных фолликул, реагирующих на давление от движения волоса.
Нервно-мышечные веретена – рецепторы растяжения поперечнополосатых мышц — нервные окончания, обладающие как чувствительной, так и двигательной иннерваций. Чувствительные нервные волокна витками обильно оплетают несколько мышечных волокон, образуя вокруг них подобие муфты.
В этой области мышечные волокна истончаются, число миофибрилл в них уменьшается, а количество ядер резко возрастает. Нервно-мышечные веретёна бывают окружены соединительнотканной капсулой.
[1] Двигательные нервные волокна образуют мелкие нервномышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус, регулируя длину волокон. Всё свободное пространство между мышечными волокнами заполнено жидкостью и ограничено тонкой капсулой.
Изменение тонуса мышцы ведет к изменению давления жидкости и передается на дендриты. Кольцеспиральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, гроздевидные окончания – только на изменение длины.
Число веретен в мышце зависит от её функции и тем выше, чем более точными движениями она обладает.[2] Нервно-сухожильные веретена – рецепторы растяжения, располагаются в месте соединения мышцы с сухожилием, веретеновидные структуры длиной 0,5-1 мм. Каждое веретено имеет капсулу из фиброцитов, которая охватывает группу сухожильных пучков, оплетенных окончаниями нервных волокон. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.[2]
- От рецептора импульс движется по рефлекторной дуге через спинной мозг и ствол мозга к ядрам таламуса и далее к коре.
- 4. Рабочие характеристики
- Количество и плотность размещения рецепторов:
— Общее количество тактильных рецепторов оценивается в 10 миллионов штук, сгруппированных в 1 миллион афферентов. — Количество свободных афферентных окончаний в коже: 50% от общего количества кожных афферентов.
— Плотность размещения свободных нервных окончаний в коже: 170 шт/см2 [4]
Размеры и прочие структурные параметры:
— Тельца Мейснера: 50-140 мкм — Колбы Крауза: 40-150 мкм — Тельца Руффини: 1-2 мм — Тельца Фатера-Пачини: 0.5-5 мм [4] — Нервно-сухожильные веретёна: — Размеры: 0.5-1 мм — Количество мышечных волокон в веретене: 15 штук — Нервно-мышечные веретёна — Количество мышечных волокон в веретене: 1-8 штук
— Диаметр выходящих нервных волокон: 17 мкм (первичные) и 8 мкм (вторичные)[5]
Скорость передачи:
- — Психофизическая оценка ёмкости тактильных каналов: 5 бит/c — Скорость передачи импульсов из свободных нервных окончаний: 1 м/c (С-волокна, IV тип)
- — Скорость передачи импульсов из инкапсулированных рецепторов: 50 м/c (А-волокна, II тип) [3]
Разрешающая способность и пороги ощущений:
- — Порог ощущения кожных рецепторов при надавливании: 10 мкм — Одновременный пространственный порог кожных рецепторов: — На губах и кончиках пальцев: 1-3 мм — На спине, плечах, бёдрах: 50-100 мм — Последовательный пространственный порог кожных рецепторов: — На губах и кончиках пальцев: 1 мм — На спине, плечах, бёдрах: 10-20 мм — Порог ощущения вибрации: 150-300 Гц
- — Минимальная ощущаемая амплитуда вибрации: 1 мкм
Модальность (специфичность к воздействиям):
— Тельца Руффини и колбы Крауза помимо механических воздействий детектируют температуру, т.е. полимодальны. А тельца Мейснера могут ощущать вибрацию. Также полимодальны неинкапсулированные свободные афференты. — Остальные рецепторы унимодальны.
Рецепторные поля (площадь рецепции):
— Для тактильных дисков площадь равна анатомическим размерам. — Для дисков Меркеля это агломерация из 30-50 дисков, обслуживаемых одним нервным волокном.
— Для телец Фатера-Паччини эта область больше анатомически размером.[4]
Адаптация (см. Таблица 1).
Таблица 1. Классификация кожных механорецепторов по скорости адаптации и адекватным стимулам.[4]
5. Регуляция Информация от рецепторов передается в центральную нервную систему через спинальные или черепномозговые нервы.
Ответвления аксонов, передающие сенсорную информацию, формируют синаптические окончания на нейронах спинного мозга (или ствола мозга в случае черепномозговых червов), затем направляются в высшие отделы нервной системы (стволовые ядра, которые в свою очередь передают информацию выше).
В каждом из этих отделов нервной системы поток сенсорной информации может фильтроваться — акцентрироваться или, наоборот, блокироваться.
Сенсорные системы построены по иерархическому принципу: сигналы от рецепторов поступают в низшие уровни центральной нервной системы (спинной мозг или ствол мозга), откуда передаются в более высшие отделы (ядра таламуса, кора больших полушарий, базальные ганглии). На каждой из этих последовательных стадий сенсорная информация преобразуется, происходит ее фильтрация.
Поток информации не односторонний, так как высшие отделы иерархии посылают сигналы в низшие отделы. Кроме того, сенсорная информация обрабатывается не цепочкой последовательных структур, а, скорее, многими областями мозга одновременно (или, как говорят, параллельно).
Параллельная обработка сенсорной информации становится очевидна в высших областях мозга, таких как кора полушарий. Здесь отдельные области специализируются на обработке отдельных элементов информации. Сенсорная информация различного рода не обрабатывается отдельно.
Во многих областях мозга, называемых ассоциативными, происходит смешение модальностей — например, в париетальной коре и буграх четверохолмия. Нейроны в этих отделах мозга реагируют на различные стимулы, например, зрительные, тактильные и слуховые.
Структурами, контролирующими сенсорную информацию, являются кора больших полушарий (в частности, префронтальная кора, играющая важную роль в контроле внимания), базальные ганглии, ретикулярная формация, таламус (в частности, ретикулярное ядро таламуса) и другие структуры. Одним из основных механизмом фильтрации сенсорной информации является торможение, которое производят ГАМК-эргические синапсы. Как правило, нейрон передающий сенсорную информацию, является возбуждающим. Фильтруют эти возбуждающие сигналы тормозные нейроны. Торможение может быть пресинаптическим (то есть блокирующим передачу сигналов по сенсорному аксону к какому-либо нейрону) либо постсинаптическим (гиперполяризующий нейрон, принимаюший сенсорные сигналы). Постсинаптическое торможение позволяет блокировать сигналы избирательно, так как принимающая клетка остается способной отвечать на другие, незаблокированные воздействия.
- На фильтрацию сенсорных сигналов также оказывают влияние такие нейромедиаторы как ацетилхолин, дофамин, эндорфины и другие.
- 6. Энергозависимость
Так как весь анализатор механорецепции является производным нервной ткани, то энергозависимость особенно высокая. Нейроны снабжаются питательными веществами и кислородом через нейроглию и расположенные в ней кровеносные сосуды.
При гипоксии и тромбах может нарушаться питание отдельных участков (например кожи или органов), начнётся ретроградная дегенерация нейронов (в некоторых случаях возможно восстановление функций волокна через некоторое время после восстановления питания) и утрату рецепции на участке.
Однако гипоксия в мозге может привести к гораздо худшим последствиям, утратой функций целых отделов или полей коры мозга.
7. Технический аналог
Аналогом механорецепторов являются устройства, основанные на пьезоэффекте. Тельца Руффини в своём составе имеют коллагеновые волокна, а они проявляют пьезоэлектрические свойства. Пьезоэлектрический эффект (пьезоэффект) состоит в том, что при механических деформации некоторых кристаллов в определённых направлениях на их гранях появляются электрические заряды противоположных знаков, т.е.
механические воздействия трансформируются в напряжение. Получается, что тельца Руффини функционируют как своеобразные пьезоэлектрические устройства, поскольку в них между нервной терминалью и коллагеновым волокном нет какой-либо иной ткани. В технике часто используется обратный пьезоэффект, когда электрический ток преобразуется в механические колебания – например, для генерации ультразвука.
Список литературы
1. Шубникова Е.А., Функциональная морфология тканей. – М: Издательство Московского университета. – 1981. – 326 с. 2. Фенькина Р.П., Дегтярев В.П., Коротич В.А., Учебное пособие по нормальной физиологии. – М:МГМСУ. – 1994г 3. Сандаков Д.Б., Курс лекций по физиологии. – Минск: биофак БГУ.
4. Шмидт Р., Основы сенсорной физиологии. – М:Мир. – 184. – 287 с.
5. Кузнецов С.Л., Мушкамбаров Н.Н., Горячкина В.Л. Руководство-атлас по гистологии, цитологии и эмбриологии.
Механорецептор — Mechanoreceptor
А механорецептор, также называемый механоцептор, это сенсорный ячейка, которая реагирует на механическое давление или искажение.
Существует четыре основных типа механорецепторов у голых или голых, млекопитающее кожа: ламеллярные тельца (Тельца Пачини), тактильные тельца (Тельца Мейснера), Нервные окончания Меркель, и луковичные тельца (Тельце Руффини).
Есть также механорецепторы в волосистой коже и волосковые клетки в рецепторах приматов, таких как макаки резус и другие млекопитающие похожи на млекопитающих и также изучались анатомически и нейрофизиологически еще в начале 20 века.[1]
Механорецепторы беспозвоночных включают: колоколообразная сенсилла и разрез сенсиллы, среди прочего.
Механорецепторы также присутствуют в клетках растений, где они играют важную роль в нормальном росте, развитии и восприятии окружающей среды.[2]
Механизм ощущения
В соматосенсорный трансдукция, то афферентные нейроны передавать сообщения через синапсы в ядра спинного столба, куда нейроны второго порядка отправить сигнал в таламус и синапс с нейроны третьего порядка в вентробазальный комплекс. Затем нейроны третьего порядка посылают сигнал в соматосенсорная кора.
Обратная связь
Более поздние работы расширили роль кожных механорецепторов для обратной связи в мелкая моторика.[3] Потенциалы одиночного действия от Тельце Мейснера, Тельце Пачини и Окончание Руффини афференты напрямую связаны с активацией мышц, тогда как Клетка Меркель-нейритный комплекс активация не вызывает мышечной активности.[4]
Типы
В голой (безволосой) коже существует четыре основных типа механорецепторов, каждый из которых имеет форму в соответствии со своей функцией. В тактильные тельца (также известные как тельца Мейснера) реагируют на легкое прикосновение и быстро адаптируются к изменениям текстуры (колебания около 50 Гц). В луковичные тельца (также известные как окончания Руффини) обнаруживают напряжение глубоко в коже и фасция. В Нервные окончания Меркель (также известные как диски Меркеля) обнаруживают устойчивое давление. В ламеллярные тельца (также известные как тельца Пачини) в коже и фасции обнаруживают быстрые колебания (около 200–300 Гц).
Рецепторы в волосяные фолликулы так называемые корневые сплетения волос[5] смысл, когда волосы меняет положение. Действительно, наиболее чувствительными механорецепторами у человека являются волосковые клетки в улитка из внутреннее ухо (никакого отношения к фолликулярным рецепторам — они названы в честь механосенсорной стереоцилии они обладают); эти рецепторы преобразовывать звук для мозга.
Механосенсорный свободные нервные окончания обнаруживать прикосновение, давление, растяжение, а также ощущение щекотки и зуда. Ощущения зуда вызваны раздражением свободных нервных окончаний химическими веществами.[5]
Барорецепторы являются типом механорецепторных сенсорных нейронов, которые возбуждаются растяжением кровеносного сосуда.
Кожный
Кожные механорецепторы реагируют на механические стимулы, возникающие в результате физического взаимодействия, включая давление и вибрацию. Они расположены в коже, как и другие кожные рецепторы.
Все они иннервируются Волокна Aβ, кроме механоприемника свободные нервные окончания, которые иннервируются Волокна Aδ. Кожные механорецепторы можно классифицировать по морфологии, по типу ощущений, которые они воспринимают, и по скорости адаптации.
Кроме того, у каждого свой рецептивное поле.
- В Медленно адаптирующийся механорецептор типа 1 (SA1), с Меркель корпускулярный орган-мишень, лежит в основе восприятия формы и шероховатости на коже.[6] У них небольшие рецептивные поля, и они вызывают устойчивую реакцию на статическую стимуляцию.
- В Медленно адаптирующиеся механорецепторы типа 2 (SA2), с Тельце-мишень Руффини, реагируют на растяжение кожи, но не были тесно связаны ни с проприоцептивной, ни с механорецептивной ролями в восприятии.[7] Они также вызывают устойчивые реакции на статическую стимуляцию, но имеют большие рецептивные поля.
- В Быстрая адаптация (RA) или Тельце-мишень Мейснера механорецептор лежит в основе восприятия трепета[8] и скользить по коже.[9] У них небольшие рецептивные поля, и они вызывают кратковременные реакции на начало и прекращение стимуляции.
- В Тельца Пачини или тельца Фатера-Пачини или ламеллярные тельца[10] лежат в основе восприятия высокочастотной вибрации.[8][11] Они также производят кратковременные ответы, но имеют большие рецептивные поля.
По ощущениям
| Эта секция нуждается в расширении. Вы можете помочь добавляя к этому. (Декабрь 2016 г.) |
По скорости адаптации
Кожные механорецепторы также можно разделить на категории в зависимости от степени их адаптации. Когда механорецептор получает стимул, он начинает выдавать импульсы или потенциалы действия на повышенной частоте (чем сильнее раздражитель, тем выше частота). Клетка, однако, скоро «адаптируется» к постоянному или статическому стимулу, и частота импульсов снизится до нормальной.
Рецепторы, которые быстро адаптируются (т.е. быстро возвращаются к нормальной частоте пульса), называются «фазовыми». Те рецепторы, которые медленно возвращаются к своей нормальной скорости активации, называются тоник.
Фазические механорецепторы полезны для восприятия таких вещей, как текстура или вибрации, тогда как тонические рецепторы полезны для измерения температуры и проприоцепция среди прочего.
Рецептивное поле
Кожные механорецепторы с небольшими точными рецептивные поля находятся в областях, требующих аккуратного воздействия (например, кончики пальцев). В кончиках пальцев и губ значительно повышена плотность иннервации медленно адаптирующихся механорецепторов I типа и быстро адаптирующихся механорецепторов I типа.
Эти два типа механорецепторов имеют небольшие дискретные рецептивные поля и, как полагают, лежат в основе наиболее низкопорогового использования пальцев при оценке текстуры, скольжения поверхности и флаттера.
Механорецепторы, обнаруженные в областях тела с меньшей тактильной остротой, имеют тенденцию к увеличению рецептивные поля.
Другие
Другие механорецепторы, кроме кожных, включают волосковые клетки, которые Рецепторы чувств в вестибулярный аппарат из внутреннее ухо, где они вносят свой вклад в слуховая система и равновесие. Кроме того, механорецепторы помогают Dionaea muscipula Ellis (Venus Fly Trap) в захвате значительных[12] эффективно добывать.[13]
Это также Юкстакапиллярные (J) рецепторы, которые реагируют на такие события, как отек легких, легочная эмболия, пневмония, и баротравма.
Связочный
Существует четыре типа механорецепторов, встроенных в связки. Поскольку все эти типы механорецепторов миелинизированы, они могут быстро передавать сенсорную информацию о положении суставов в центральную нервную систему.[14]
- Тип I: (маленький) Низкий порог, медленная адаптация как в статических, так и в динамических настройках
- Тип II: (средний) Низкий порог, быстро адаптируется к динамическим настройкам
- Тип III: (большой) Высокий порог, медленно адаптируется к динамическим настройкам
- Тип IV: (очень маленький) Высокопороговые болевые рецепторы, сообщающие о травме.
В частности, считается, что механорецепторы типа II и III связаны с чувством проприоцепция.
Ламеллярное тельце
Пластинчатые тельца, или тельца Пачини, представляют собой рецепторы давления, расположенные в коже, а также в различных внутренних органах. Каждый связан с сенсорным нейроном.
Из-за его относительно большого размера можно изолировать одиночное ламеллярное тельце и изучить его свойства.
Механическое давление различной силы и частоты может быть приложено к тельце с помощью иглы, и результирующая электрическая активность будет обнаружена с помощью электродов, прикрепленных к препарату.
Деформация тельца создает в сенсорном нейроне, возникающем внутри него, генераторный потенциал. Это ступенчатая реакция: чем больше деформация, тем больше потенциал генератора. Если потенциал генератора достигает порогового значения, срабатывает залп потенциалов действия (нервных импульсов). узел Ранвье сенсорного нейрона.
Как только порог достигнут, величина стимула кодируется в частоте импульсов, генерируемых нейроном. Таким образом, чем сильнее или быстрее деформация отдельного тельца, тем выше частота нервных импульсов, генерируемых в его нейроне.
Оптимальная чувствительность ламеллярного тельца составляет 250 Гц, частотный диапазон, создаваемый на кончиках пальцев текстурами, состоящими из элементов менее 200микрометры.[15]
является
Мышечные веретена и рефлекс растяжения
В коленный рефлекс широко известный рефлекс растяжения (непроизвольный удар голени), вызванный ударом по колену молотком с резиновой головкой. Молот бьет сухожилие который вставки ан разгибатель мышца передней части бедра в голень. Постукивание по сухожилию растягивает мышцу бедра, что активирует рецепторы растяжения внутри мышцы, называемой мышечные веретена.
Каждое мышечное веретено состоит из сенсорных нервных окончаний, обернутых вокруг специальных мышечных волокон, называемых волокна веретена (также называемые интрафузальными волокнами). Растягивание волокна веретена вызывает в сенсорном нейроне залп импульсов ( Я нейрон), прикрепленный к нему.
Импульсы проходят по сенсорному аксону к спинному мозгу, где они образуют несколько видов синапсов:
- Некоторые из ветвей синапсов аксонов I-a непосредственно с альфа двигательные нейроныОни передают импульсы обратно к той же мышце, заставляя ее сокращаться. Нога выпрямляется.
- Некоторые ветви синапсов I-a аксонов с тормозящими интернейронами в спинном мозге. Это, в свою очередь, синапс с двигательными нейронами, ведущими обратно к антагонистической мышце, сгибателю задней части бедра. Блокируя сгибатель, эти интернейроны способствуют сокращению разгибателя.
- Есть и другие ветви синапса аксонов I-a с интернейронами, ведущими к центрам мозга, например мозжечку, которые координируют движения тела.[16]
Смотрите также
Примечания
- ^ Адриан Э.Д., Умрат К. (октябрь 1929 г.). «Импульсный разряд из тельца пациента». Журнал физиологии. 68 (2): 139–54. Дои:10.1113 / jphysiol.1929.sp002601. ЧВК 1402853. PMID 16994055.
- ^ Monshausen, Gabriele B .; Хасуэлл, Элизабет С. (декабрь 2013 г.). «Сила природы: молекулярные механизмы механовосприятия у растений».
Журнал экспериментальной ботаники. 64 (15): 4663–4680. Дои:10.1093 / jxb / ert204. ISSN 0022-0957. ЧВК 3817949. PMID 23913953.
- ^ Йоханссон Р.С., Фланаган-младший (май 2009 г.). «Кодирование и использование тактильных сигналов от кончиков пальцев в задачах манипулирования объектами» (PDF). Обзоры природы. Неврология. 10 (5): 345–59.
Дои:10.1038 / номер 2621. PMID 19352402. S2CID 17298704.
- ^ McNulty PA, Macefield VG (декабрь 2001 г.). «Модуляция текущей ЭМГ различными классами низкопороговых механорецепторов в руке человека». Журнал физиологии. 537 (Pt 3): 1021–32. Дои:10.1111 / j.1469-7793.2001.01021.x. ЧВК 2278990. PMID 11744774.
- ^ а б Тортора, Джерард Дж., Автор. Основы анатомии и физиологии. ISBN 978-0-7303-5500-7. OCLC 1059417106.CS1 maint: несколько имен: список авторов (связь)
- ^ Джонсон К.О., Сяо СС (1992). «Нейронные механизмы тактильного восприятия формы и текстуры». Ежегодный обзор нейробиологии. 15: 227–50. Дои:10.
1146 / annurev.ne.15.030192.001303. PMID 1575442.
- ^ Torebjörk HE, Очоа JL (декабрь 1980 г.). «Специфические ощущения, вызванные активностью отдельных идентифицированных сенсорных единиц в человеке». Acta Physiologica Scandinavica. 110 (4): 445–7. Дои:10.1111 / j.1748-1716.1980.tb06695.x. PMID 7234450.
- ^ а б Talbot WH, Darian-Smith I, Kornhuber HH, Mountcastle VB (март 1968 г.). «Чувство трепетания-вибрации: сравнение возможностей человека с паттернами реакции механорецептивных афферентов от руки обезьяны». Журнал нейрофизиологии. 31 (2): 301–34. Дои:10.1152 / jn.1968.31.2.301. PMID 4972033.
- ^ Йоханссон RS, Вестлинг G (1987). «Сигналы тактильных афферентов от пальцев, вызывающие адаптивные двигательные реакции во время точного захвата». Экспериментальное исследование мозга. 66 (1): 141–54. Дои:10.1007 / bf00236210. PMID 3582528. S2CID 22450227.
- ^ Бисвас А., Маниваннан М., Сринивасан М.А. (2015).
«Многоуровневая многослойная биомеханическая модель тельца пациента». Транзакции IEEE по тактильности. 8 (1): 31–42. Дои:10.1109 / TOH.2014.2369416. PMID 25398182. S2CID 24658742.
- ^ Бисвас А., Маниваннан М., Сринивасан М.А. (2015). «Порог вибротактильной чувствительности: нелинейная стохастическая механотрансдукционная модель тельца Пачини».
Транзакции IEEE по тактильности. 8 (1): 102–13. Дои:10.1109 / TOH.2014.2369422. PMID 25398183. S2CID 15326972.
- ^ Чамовиц Д. (2012). Что знает растение: полевой путеводитель по чувствам (1-е изд.). Нью-Йорк: Scientific American / Farrar, Straus and Giroux. ISBN 9780374533885. OCLC 755641050.
- ^ Волков А.Г., Форд-Такетт В.
, Волкова М.И., Маркин В.С. (2014-02-10). «Морфирующиеся структуры Dionaea muscipula Ellis во время открытия и закрытия ловушки». Сигнализация и поведение растений. 9 (2): e27793. Дои:10.4161 / psb.27793. ЧВК 4091236. PMID 24618927.
- ^ Майкельсон Дж. Д., Хатчинс С. (март 1995 г.). «Механорецепторы связок голеностопного сустава человека».
Журнал костной и суставной хирургии. Британский том. 77 (2): 219–24. Дои:10.1302 / 0301-620X.77B2.7706334. PMID 7706334.
- ^ Scheibert J, Leurent S, Prevost A, Debrégeas G (март 2009 г.). «Роль отпечатков пальцев в кодировании тактильной информации, полученной с помощью биомиметического датчика». Наука. 323 (5920): 1503–6. arXiv:0911.4885.
Bibcode:2009Sci … 323.1503S. Дои:10.1126 / science.1166467. PMID 19179493. S2CID 14459552.
- ^ Кимбалл JW (2011). «Механорецепторы». Страницы биологии Кимбалла. Архивировано из оригинал 27 февраля 2011 г.